
Brussels – European Parliament – Dec 08, 2003
How can we best help Africa? How can we set priorities aimed at bringing under
control what is described as an AIDS epidemic? For twenty years, all AIDS research
has been based on the HIV hypothesis. Do we now have reasons to question this
hypothesis? Yes, because there is a major problem with isolation and purification
of HIV. The major problem being that, in spite of innumerable claims to the
contrary, this retrovirus has never been isolated nor purified in a scientifically
acceptable manner that would satisfy the classic requirements of virology.
To demonstrate the problem’s magnitude it appears necessary to compare current results on HIV with those obtained, previously, in experimental pathology, on another retrovirus known to be significantly associated with a particular leukaemia of laboratory mice, the Friend leukaemia. Both retroviruses, i.e. the Friend leukaemia virus, and the HIV hypothetically related to AIDS, share extremely similar morphology under the electron microscope, have identical diameters, and sediment at the same density in sucrose gradients. A direct comparison between isolating and purifying these two different retroviruses is, therefore, entirely appropriate.
Mice suffering from the Friend leukaemia have considerable numbers of retroviral particles in their blood stream. This phenomenon, described as “Viraemia“ in the past (1), would be called “Viral load“, in today ’s terminology. From only a few ml of the blood plasma of leukaemic mice, the viral particles could be readily separated by a simple technique of ultrafiltration, then sedimented by high-speed centrifugation and finally prepared for electron microscopy. The results are illustrated in the first slide.
On this electron microscope image, a uniform population of virus particles is clearly recognized. They all have the same diameter and morphology, and one has to look very carefully to identify rare, non-viral structures, attesting to the high degree of purification of these retroviral particles. Such samples of purified retrovirus were successfully used to identify viral proteins and to extract viral RNA. The method applied to achieve this purification of a typical retrovirus is rapid, inexpensive and highly reproducible.
Most surprisingly, nobody has ever succeeded in demonstrating HIV particles in the blood of any AIDS patient by this simple method, even though patients were selected for presenting a so-called high “Viral load” as determined by PCR methods. This embarrassing lack of electron microscope evidence for substantiating the nature of the so-called viral load in AIDS patients was first reported during an important AIDS conference that took place in Pretoria, S.A., in May 2000 (2). None of the AIDS experts present at that conference could demonstrate the presence of retroviral particles in the blood of AIDS patients. Moreover, almost two years ago, a substantial award ($100.000) was officially offered (3) to anybody who would demonstrate HIV particles in the blood of allegedly high viral load patients. Two years later, the award has still not been claimed. Obviously, what was so readily and reproducibly demonstrated in leukaemic mice has never been observed in any AIDS patient.
ALLEGED CLAIMS OF HIV ISOLATION BASED ON NON-SPECIFIC “MARKERS”.
Over the past 20 years, the medical literature has been inundated with innumerable papers, attempting to dodge the lack of electron microscope evidence for the presence of retroviral particles in samples directly collected from AIDS patients. In all these papers, the missing retroviral particles have been swiftly substituted with so-called HIV “Markers”. These “Markers” were of physical, biochemical, or genetic nature.
Physical markers.
As known for a very long time, classic retroviruses identified in chicken, mice and cats, all share the same shape and density, and therefore sediment at exactly the same level during high-speed centrifugation in sucrose gradients. Actually, they all sediment at the density of 1.16 gm of sucrose per ml (4). The alleged HIV being classified as a retrovirus, it was logical to expect it to sediment at that same density.
However, as was also well known years before the emergence of AIDS, a large variety of cellular fragments and debris also sediment at that density (5, 6). Collecting material sedimenting at that density does not, therefore, demonstrate the isolation of retroviruses, unless a careful control by electron microscopy rules out any contamination by cellular debris. The importance of these essential controls was stressed during a conference that took place in Paris, in 1973 (4). Most surprisingly, in the same laboratory of the Pasteur Institute, then years later, in 1983, a paper was published (7), in which these controls were missing. It appeared later (20), however, that these controls were attempted but gave discouraging results. Still, in the title of that paper, “isolation” of a new retrovirus, the future HIV, was announced. Dramatically enough, this is the paper that placed AIDS research on highly questionable tracks for the following two decades.
Biological markers.
In 1970, Temin (8) and Baltimore (9) discovered the activity of a so far totally unexpected enzyme in allegedly purified samples of experimental retroviruses. This enzyme was called “Reverse transcriptase” because it induces DNA synthesis from RNA templates. It was indeed a fundamental discovery that revolutionized molecular biology. This enzyme activity was first observed in RNA tumour viruses and was, therefore, initially thought to represent a characteristic “marker” of these viruses which, consequently, received a new name: “Retroviruses”. Ever since, reverse transcriptase activity has been used as a “marker” for HIV…
However, shortly after the publications by Temin and Baltimore, it was discovered that reverse transcriptase activity was not restricted to “retroviruses” but was in fact a most common phenomenon in biology (10, 11), as reviewed by Varmus in 1987 (12). Unfortunately, and yet again, Temin and Baltimore omitted to verify the purity of the viral samples on which their observations were made. Consequently, any contamination of these samples by cell, bacterial, or mycoplasma debris could just as well have explained the presence of the reverse transcriptase activity observed by these authors. In 1983, the Pasteur group based their claim for the isolation of a new retrovirus primarily on 1) the detection of reverse transcriptase activity in 2) material sedimenting at the density of 1.16 gm/ml. These two criteria completely lose significance if the data is not supported by electron microscopy verification, excluding the interference of non-viral contaminants known to be frequently present in allegedly “purified” retroviruses (5, 6).
Several proteins, allegedly of viral origin, are frequently used as “specific” HIV markers, p24 for example. Doubts concerning its specificity have been expressed for a long time (15). The complete lack of agreement between results obtained with p24 and measurements of “viral load” obtained by PCR were recently stressed (13). Surprisingly, in Western blot tests, 40% of …dogs reacted positively with proteins obtained by genetic recombination technology, such as gp120, gp47, p31 and p24 (14). This had to be expected since Eleni Papadopoulos, Val Turner and the Perth Group had initially, extensively demonstrated the total lack of specificity of all the alleged HIV structural proteins in a paper, published 10 years ago in Nature/biotechnology (15), a fundamental paper that was completely ignored. To cite only key examples, gp41 probably corresponds to actin, and gp120-160 are likely oligomers of gp41. Evidently, cell debris contaminating very poorly purified retroviral samples may also readily account for the presence of alleged retroviral markers, and frequently reported “successes” in HIV “isolation” most likely result from abusive reliances on non-specific “markers”.
Genetic markers and measurements of “viral load”.
This approach could seem more attractive for two reasons: 1) it applies directly to a patient’s blood, therefore avoiding all the uncertainties of complex cell cultures, and 2) it is supposed to be a quantitative method.
However, as already stressed, it has never been possible to visualize any retroviral particle by electron microscopy in the blood of AIDS patients, even though these patients are selected for having a so-called very high “viral load” (2). Moreover, it appears very likely that PCR methods amplify small RNA fragments, more frequently observed under conditions of stress and of chronic illnesses (16), and which include retroviral segments originating from human endogenous retrovirus. This is not surprising since about 2% of the human genome have marked homology with the retroviral genome (17). Consequently, “measuring” the “viral load” by PCR methods is likely to have no relationship whatsoever with real quantification of a hypothetical exogenous (*) HIV viremia. Kary Mullis himself, Nobel Prize laureate for his discovery of the PCR method, categorically rejects the use of “his” method for quantitative measurements of a hypothetical HIV viremia (18).
The abuse of… beautiful pictures.
The “viral load” of newspapers and magazines, all over the world is extremely high, meaning the number of pictures of HIV published almost daily in the world’s press. These pictures are extremely attractive, and are frequently rich in artificial colors. They clearly exemplify the danger of misinforming the public with computer graphics. To publish such images brings to the attention of the general public, and of the medical profession as well, an apparently crystal-clear message: “Yes, HIV has been isolated since one can portray it under the electron microscope”.
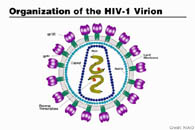
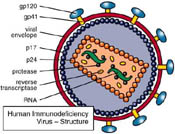
All these images represent computerized rationalizations and embellishments of actual electron microscope pictures similar to those illustrating, for example, Barré-Sinoussi’s paper (7). But not one of these pictures originated directly from one single AIDS patient! They ALL originated from complex cell cultures prepared in various laboratories (19), cultures that have been described as “real retroviral soups”…(20). Indeed, everything was done to make sure that retroviral particles (and the celebrated budding forms) would appear in these cultures. What was not done were the essential verification experiments that could have clarified the endogenous (**) origin of these viruses. Even if these control experiments had been done, their results were apparently never published. We are still waiting for a newspaper that would publish beautiful computer graphics of HIV and would have the honesty to explain to their readers that all these still have to be confirmed with samples originating directly from AIDS patients…
In AIDS research, most of the cell cultures used are mixed and hyper-stimulated.
Mixed, because they contain, for example, lymphocytes from a patient plus the H9 cells from Gallo’s lab, cells well known as chronic carriers of retroviruses (21). Or, as in the Pasteur Institute case (7), lymphocytes from an AIDS suspected patient plus lymphocytes isolated from umbilical cord blood that originated in the placenta and, as known since 1979 (22), are likely to carry human endogenous (**) retrovirus.
These cultures are hyperstimulated with one or two growth factors such as phytohemagglutinin (PHA), T cell lymphocyte growth factor (TCGF), or interleukin2, or corticosteroid hormones. All these factors are known to activate the expression of endogenous retroviruses (HERVs) that are defective viruses and may acquire envelopes and bud on the surface of cells activated by these factors. Presumably, this is exactly what happened when cord blood lymphocytes were activated with PHA and TCGF in the Pasteur 1983 experiments (7). Unfortunately, the control experiments needed to verify this interpretation remain to be done.
In short, it seems that electron microscopy was not used when it was essential to demonstrate the absence of contaminating cell debris in allegedly purified virus preparations, and misinterpreted when stimulated cord blood lymphocytes showed budding retroviruses.
Conclusion.
Indeed, HIV has never been properly isolated, nor purified, and, consequently, the HIV/AIDS hypothesis has to be fundamentally reappraised (23, 24, 25, 30, 32).
More precisely, without HIV purification HIV specific antigens could never have been rigorously identified (15). Still, so-called HIV antigens are instrumental in all the serological tests allegedly identifying specific HIV antibodies, ELISA, Western blot, and more recent rapid tests such as « Capillus », « Determine », and « Vironostika ». Recombinant DNA technology for « viral » antigens gives certainly purer products, but fails to confer the missing specificity. No surprise, therefore, that dozens of different medical conditions, including tuberculosis, malaria, leprosy, multiple blood transfusions, many vaccines, multiparity, etc. all give false-positive « HIV » tests (26).
Retroviral particles have unquestionably been observed, not directly in AIDS patients, but in mixed, hyper-stimulated cell cultures (7). They most likely represent forced expression, in cell cultures, of human endogenous retroviruses (17), the hypothetical role of which, in the etiology of AIDS, has never been proved.
The HIV particles, missing from the patients, were conveniently substituted by molecular « markers », because the HIV=AIDS hypothesis had to be saved at all cost ( see the Durban Declaration, 27), even at the price of scientific integrity (28).
If AIDS were indeed caused by a retrovirus, how can we explain that 20 years of considerable research efforts, based exclusively on that single hypothesis, have failed to isolate the responsible exogenous retrovirus? Twenty years to end up with no curative treatment, no vaccine, and no verifiable epidemiological predictions…
Obviously, time is pressing us to ask courageously the essential question i.e. is the HIV=AIDS hypothesis correct? Because it is possible to view AIDS differently, outside the field of infectious diseases, and outside the field of retrovirology (29). And in this perspective, that is loaded with optimistic predictions, all the difficulties encountered in attempted isolation and purification of the hypothetical HIV may find an extremely rational explanation. Indeed, doubts concerning the very existence of HIV are nothing new, and were expressed by several dissident scientists several years ago (30, 31). I completely share these doubts. Let us not forget the title of Peter Duesberg’s book (33) published in 1996 : “Inventing the AIDS Virus”…
Consequently, priorities for medical assistance to sub-saharan Africa should, most urgently, be revised as follows :
1) Treat all endemic tropical diseases with their specific treatments.
2) Stop all use of antiretroviral drugs until the isolation of HIV and its pathogenicity are scientifically established.
3) Stop using highly crossreacting serological tests, the HIV specificity of which is far from demonstrated.
4) Provide African people with means do combat malnutrition, clean drinking water, proper housing and sanitation, and efficient health-care infrastructures.
Thank you.
Footnote : (*)(**) Exogenous, i.e. having a cause external to the body, contrarily Endogenous i.e. originating within the body.
1) de Harven E. Viremia in Friend murine leukemia : the electron microscope approach of the problem. Pathologie-Biologie 1965; 13 :125-134. See also : de Harven E., Pioneer deplores « HIV », Continuum 1997, vol 5 n°2, page 24.
2) de Harven E. Summary statement. Interim Report of the Aids Advisory Panel, Pretoria, SA, May 2000. Published by the South Africa Government, on April 4, 2001.
3) RusselA..http://www.redflagsweekly.com/Thursdayreport/prize.html
4) Sinoussi F, et al. Purification and partial differentiation of the particles of murine sarcoma virus (M.MSV) according to their sedimentation rates in sucrose density gradients. Spectra, N°4, 1973, pp 239-243.
5) Bess JW et al. Microvesicles are a source of contaminating cellular proteins found in purified HIV-1 preparations. Virology 1997; 203 ;134-144.
6) Gluschankof P. et al. Cell membrane vesicles are a major contaminant of gradient-enriched human immuodeficiency virus type-1 preparations. Virology 1997; 230: 125-133.
7) Barré-Sinoussi F. et al. Isolation of a T-lymphotropic retrovirus from a patient at risk for asquired immune deficiency syndrome (AIDS). Science 1983; 220: 868-871.
8) Temin HM, Mizutani S. RNA-dependent DNA polymerase in viriona of Rous sarcoma virus. Nature 1970; 226: 1211-1213.
9) Baltimore D. RNA-dependant DNA polymerase. Nature 1970: 1209-1211.
10) Ross J et al. Separation murine cellular and murine leukemia virus DNA polymerases. Nature New Biology 1971 ; 231 :163-167.
11) Beljanski M. Synthèse in vitro de l’ADN sur une matrice d’ARN par une transcriptase d’Esscherichia coli. C.R. Acad. Sci 1972 ; 274 :2801-2804.
12) Varmus H. Reverse transcription. Sci. Am. 1987 ;257 :48-54.
13) Franchi F. In search of HIV ; http://www.cesil.com/0898/en frah08.htm
14) Strandstrom HV et al. Studies with canine sera that contain antibodies which recognize human immunodeficiency virus structural proteins. Cancer Research 1990 ;50 :5628s-5630s.
15) Papadopulos-Eleopulos E et al. Is a positive Western blot proof of HIV infection ? Bio/Technology 1993 ;11 :696-707.
16) Urnovitz HB et al. RNAs in the sera of Persian Gulf War veterans have segments homologous to chromosome 22Q11.2. Clin Diagn Lab Immunol 1999 ;6/3 :330-335. See also http://www.chronicillnet.org.
17) Löwer R et al. The viruses in all of us : characteristics and biological significance of human endogenous retrovirus sequences. Proc Natl Acad Sci USA 1996 ;93 :5177-5184.
18) Mullis K. « Dancing naked in the Mine Field ». Pantheon, 1998.
19) Gelderblom HR. HIV sequence data base : fine structure of HIV and SIV. http://hivweb.lanl.gov/HTML/reviews/Gelderblom.html.
20) Tahi D. Did Montagnier discover HIV ? « I repeat, we did not purify ! ». Continuum 1997 ;5 :30-34.
21) Dourmashkin RR et al. The presence of budding virus-like particles in human lymphoid cells used for HIV cultivation. VIIth International Conference on AIDS. Firenze 1992 :122.
22) Panem S. C type virus expression in the placenta. Curr Top Pathol 1979; 66: 175-189.
23) Shenton J. « Positively False ». I.B. Tauris & Co, publ., London, 1998.
24) Hodgkinson N. « The Failure of Contemporary Science – How a Virus that Never Was Deceived the World » . Fourth Estate Limited, publ., London 1996.
25) Russeil R. « Enquête sur le Sida – Les Vérités Muselées » Editions Vivez Soleil, publ., Chêne-Bourg/Genève, 1996.
26) Johnson C. Whose antibodies are they anyway ? Continuum Sept/Oct. 1996.
27) Weiss R, and Wain-Hobson S. The Durban declaration. Nature 2000; 406 : 15-16.
28) Stewart GT et al. Not all accepted the Durban Declaration. Nature 2000; 407 : 286.
29) Duesberg P, Köhnlein C and Rasnick D. The chemical bases of the various AIDS epidemics : recreational drugs, anti-viral chemotherapy and malnutrition. J. Biosci June 2003, 28 #4, pp 383-412. See French translation in www.altheal.org
30) Papadopulos-Eleopulos E. A brief history of retroviruses. Continuum 1997; 5 :25-29.
31) Lanka S. HIV, reality or artefact? Continuum 1995; 3, #1, 4-9.
32) Rasnick D. The AIDS Blunder. « Mail & Guardian », Johannesburg, S.A., Jaznuary 24, 2001.
33) Duesberg P. « Inventing the AIDS Virus ». Regnery Publishing, Inc., Washington, D.C. 1996.Etienne de Harven, MD
Formerly Member of the Sloan Kettring Institute, New York, NY,
Emerit. Prof. Pathology, Univ. of Toronto,
Member of President T. Mbeki AIDS Advisory Panel.Address for correspondence :
« Le Mas Pitou », 2879 Route de Grasse, 06530 Saint Cézaire, France.
E -mail <pitou.deharven@wanadoo.fr>
Tel or Fax (33) 4 93 60 28 39.
Back to Isolation General index